Nacionalinės mokslo programos " Agro-, miško ir vandens ekosistemų tvarumas" projektą
MIŠKOEKOKAITA SIT-4/2015 vykdo LAMMC Miškų instituto ir Gamtos tyrimų centro Botanikos instituto
bei Lenkijos Miškų tyrimo instituto (IBL) mokslininkų grupė:
A.Pliūra A., V.Suchockas V., R.Verbylaitė, G.Bajerkevičienė, J.Jankauskienė, V.Lygis, J.Labokas ir D.Dobrowolska.
Problematika.
Klimato kaitos modeliai rodo, kad iki šio amžiaus pabaigos oro temperatūra Žemėje padidės
ne mažiau kaip 2°C, o Lietuvos platumose – 4°C. Kartu dažnės klimato ekstremumai - karščio bangos,
dažnesnės ir stipresnės sausros, gausių kritulių periodai,
stiprios ankstyvos šalnos, audros, švelnios žiemos su sumažėjusia sniego danga ir įšalo gyliu ir kt.
Naujausi tyrimai rodo, jog augalai yra labiau jautrūs klimato ekstremumams
nei palaipsniams klimato pokyčiams. Tai tiesiogiai arba netiesiogiai gali sukelti medžiams stresą,
sutrikdyti jų augimo ritmą, vystymąsi, augimą ar net juos pažeisti.
Iškyla pavojus, jog medžiai, būdami ilgaamžiai organizmai, gali nesugebėti
prisitaikyti prie stresą sukeliančių klimato ekstremumų. Kadangi skirtingos medžių rūšys nevienodai jautrios
įvairiems klimato kaitos ir susijusiems stresoriams,
jautriausios gali labiausiai nukentėti, dėl ko pakistų miško ekosistemų rūšinė sudėtis ir struktūra.
Ekologiniai tyrimai
ir modeliavimai rodo, kad įvairių medžių rūšių arealų eko-klimatinis apvalkas ir miško augalijos zonos
dėl klimato kaitos sparčiai slinksis į šiaurę – 300-800 km per šimtmetį, tačiau medžių gamtinės
migracijos greitis bus ženkliai mažesnis – 10-70 km per šimtmetį, o medynuose - dar mažesnis.
Taigi, esamos miško eksosistemos bus veikiamos ne tik klimato ekstremumų, bet greitai atsidurs
skirtingose ekologinėse sąlygose, nei tose, kuriomis jos įsikūrė, formavosi ir adaptavosi.
Ypač neigiamą tiesioginį ir netiesioginį tokios kaitos poveikį gali pajusti paprastoji eglė. Jos
išplitimo arealo pietinė riba, esanti šiaurinėje Lenkijoje, neišvengiamai trauksis į šiauresnes platumas.
Todėl Lietuvoje reikėtų pradėti galvoti, kaip veisti miškus iš piečiau kilusių kilmių
(pietinės Lenkijos ir Vokietijos) ir net svetimžemes medžių rūšis (bekotį ąžuolą, hibridines
tuopas, maumedį, sitkinę eglę). Mūsų šalyje dėl didėjančios temperatūros, CO2 koncentracijos
ir gausėjančių kritulių daugeliui lapuočių medžių rūšių augimo sąlygos turėtų gerėti.
Daugelio medžių sėjinukai yra jautrūs pavasarinėms ir rudens šalnoms, kurios tampa vis dažnesnės
ir stipresnės, ypač didelėse kirtavietėse, kur mikroklimatas yra kitoks, nei miške.
Be to, žiemos su dažnėjančiais šiltais periodais
suardo augalų žiemos ramybės ciklą ir, augalams pradėjus anksčiau sprogti, šalnų pavojus dar
padidėja. Dėl to medynų atsikūrimo stadijoje atsiranda stiprus selekcinis spaudimas ir
vyksta aktyvūs selekciniai procesai. Dalies medžių (pvz., ąžuolo) žiedai taip pat
labai jautrūs šalnoms. Tad nušalus vienos ar kitos fenologijos
žiedams, palikuonių genotipinė struktūra atsikuriančiuose miškuose stipriai pakinta. Lapuočių
medžių rūšys gana jautrios gruntinio vandens lygio svyravimams, o dėl klimato ekstremumų šis
svyravimas didėja ir neigiamai įtakoja tiek jaunus, tiek ir brandžius medžius. Pakilusi
temperatūra, šiltos žiemos ir pakitęs drėgnumas sudaro palankias sąlygas epideminiam vietinių
ir net invazinių kenkėjų ir ligų išplitimui, pvz. grybo Hymenoscyphus fraxineus, sukėlusio masinę uosynų
džiūtį visoje Europoje.
Vykstantys globalūs klimato pokyčiai bei žmogaus ūkinės veiklos sukelti vietiniai aplinkos pokyčiai
vis stipriau veikia augalų populiacijas, kurios formavosi palyginus lėtai kintančiomis aplinkos
sąlygomis per daugelį šimtmečių. Dabartiniai pokyčiai žymiai spartesni nei tie, kurie vyko
stipriai keičiantis klimatui poledynmečio laikotarpiu. Toks stiprus stresinis spaudimas gali
sukelti ne tik rūšių sezoninio vystymosi ir fiziologinius pokyčius, bet per intensyvėjančią
natūraliąją atranką iššaukia ir genetinių savybių bei populiacijų genetinės struktūros pokyčius,
sumažinti genetinę įvairovę ir kartu – populiacijų tolesnės genetinės adaptacijos ir evoliucijos galimybes.
Dėl įvairių augalų rūšių, jų populiacijų ir genotipų skirtingo adaptacinio pajėgumo, keičiantis
klimatui ir aplinkos sąlygoms, gali labai pakisti ir rūšių konkurencingumas ir paplitimas,
o tai gali lemti ekosistemų struktūros, biologinės įvairovės, miško ekosistemų produktyvumo ir funkcionavimo pokyčius.
Vis dažnėjant su klimato kaita susijusių abiotinių (audros, gaisrai) ir biotinių
(kenkėjų ir ligų epidemijos) trikdžių mastams ir dažniui, iškilo realus pavojus esamų miško
ekosistemų tvarumui, mūsų hemiborealiniams miškams būdingoms sukcesijoms, pilnaverčiam miškų
atsikūrimui ir jų genetinei įvairovei. Vėjo, gaisrų ir ligų pažeistose ekosistemose ypač
sumažėja karkasinių ir susijusių augalų rūšių efektyvusis populiacijų dydis (derančių individų
skaičius) ir kartu genetinė įvairovė, kurios gali nebepakakti rūšių fiziologinei ir genetinei
adaptacijai, populiacijų tvarumui išlaikyti ir genetiškai pilnaverčiam atsikūrimui užtikrinti.
Trumpalaikis prisitaikymas gali būti pasiekiamas per fiziologinį, fenotipinį ir morfologinį
plastiškumą. Pavyzdžiui, kai medžiai nepalankiomis sąlygomis sumažina dujų apykaitą ar
išgarinimą užverdami žioteles lapuose, ar net numesdami lapus, sulėtindami augimą ar keisdami
morfologiją. Skirtingos medžių rūšys pasižymi savita fiziologine ir biochemine reakcija į streso veiksnius.
Čia dažnai pasireiškia apigenetiniai reiškiniai – adaptaciniai pokyčiai
pasikeitusios aplinkos sąlygoms ar dėl stresorių poveikio vyksta dėl genų raiškos pokyčių.
Populiacinės adaptacijos atveju dėl nepalankių sąlygų iškristų dalis mažiau atsparių genotipų
ir populiacija jau toje pačioje generacijoje taptų kiek labiau prisitaikiusi. Biocenotinės
adaptacijos atveju iš miško ekosistemose jau sekančioje kartoje gali nelikti dalies medžių
rūšių – vyks sukcesijos. Tačiau ilgalaikę genetinę adaptaciją
dideliems aplinkos pokyčiams ir net rūšies evoliuciją gali užtikrinti tik pakankamai didelė genetinė variacija
ir vykstanti gamtinė atranka. Tačiau genetinės adaptacijos procesai lėtesni – reikia vienos ir daugiau
generacijos pasikeitimo, kad dėl gamtinės atrankos iškristų menkai prisitaikę genotipai,
o geriau prisitaikę išplistų. Epigenetiškai reguliuojamas (dėl genų įjungimo ar išjungimo ar jų raiškos
pokyčių) fenogenetinis plastiškumas (genetinė variacija pagal ekologinį atsaką) yra taip pat
adaptacijai labai svarbi genetinės variacijos dalis. Genetinės variacijos pokyčiai klimato kaitos sąlygomis
skirtingoms medžių rūšims sunkiai prognozuojami. Tad genetinės variacijos pokyčių tyrimai labai aktualūs.
Įvairiose šalyse atliekama nemažai miško ekosistemų dinamikos tyrimų, kuriuose analizuojamas
įvairių gamtinių ir antropogeninių trikdžių poveikis miško ekosistemų tvarumui ir vystymuisi,
genetinei ir visai biologinei įvairovei, pabrėžiant, kad vidutinio stiprumo trikdžiai yra
būtini ekosistemų vystymuisi, tačiau stiprūs trikdžiai ekosistemas gali suardyti (Attivil 1994, Bengtsoon et al 2000,
Schulze et al 2007, Linder et al 2010, Bollmann and Braunisch 2013 ir kt.). Palyginti nedaug žinoma,
kaip atskiros medžių rūšys ir jų bendrijos sugeba adaptuotis prie besikeičiančių ar stresinių
aplinkos sąlygų, kurios jų yra geriausiai išvysčiusios streso indukuojamų pažeidimų eliminavimo
mechanizmus. Taip pat nėra atskleistas medžių fiziologinės apsaugos strategijos mechanizmas
reaguojant tiek į pavienius, tiek ir kartu veikiančius, dažnai sinergiškai – vienas kito nepalankų
poveikį stiprinančius, ar kitaip sąveikaujančius aplinkos veiksnius. Trūksta žinių ir apie tai, kaip ir kiek fenotipinis
plastiškumas, ekologinis atsakas, genetinė įvairovė ir evoliucinė adaptacija leis rūšims
prisitaikyti prie naujų aplinkos sąlygų Lietuvoje.
Ekologiniu požiūriu rizikingiausių - gamtinių ar antropogeninių trikdžių suardytų miško ekosistemų
atsikūrimas vyksta dideliuose atviruose plotuose, kurie veikiami kontrastingų temperatūrų
dienomis ir naktimis, drėgmės režimo sutrikdymo, intensyvesnės UV spinduliuotės ir padidėjusios
priežemio ozono koncentracijos. Atsikūrimo stadijoje miško ekosistemos yra jautriausios ir
lengviausiai pažeidžiamos, o jų atsikūrimo eiga iš esmės apsprendžia naujai besiformuojančios
miško bendrijos tipą, struktūrą ir būsimą tvarumą. Todėl tyrimai šioje ekositemų vystymosi
stadijoje yra ypač aktualūs ir svarbūs.
Dalies medžių rūšių (uosio, beržo, eglės) atsaką į trumpalaikį ozono ir UV spinduliuotės
poveikį tyrėme mokslo programoje APLIKOM (2010-2012), tačiau nebuvo tirtas komplekso veiksnių
poveikis derinyje su padidinta temperatūra, drėgme ir CO2 koncentracija bei su klimato kaita
susijusiais stresoriais - šalnomis, sausra, kaitra. Ekosistemų degradacija yra daugiaplanis
procesas, kur streso veiksniai veikia integruotai. Tačiau iki šiol dauguma medžių tyrimų
stresorių ar patogenų poveikyje klimatinėse kamerose Lietuvoje ir užsienyje vykdomi paprastai
tik veikiant kuriuo nors vienu stresiniu veiksniu ir tik atskiroms rūšims, todėl vykdomų
kompleksinių tyrimų naujumas neabejotinas ir sudaro geras galimybes publikuoti tyrimų rezultatus
tarptautiniuose moksliniuose žurnaluose.
Vykdomų tyrimų tikslas atitinka Nacionalinės programos tikslą - kompleksiniais mokslo tyrimais gauti,
išanalizuoti ir apibendrinti naujas mokslo žinias apie klimato kaitos ir ekosistemų išteklių
naudojimo poveikį Lietuvos ekosistemoms ir apima vienas iš svarbiausių - miško ekosistemas
jautriausiame jų formavimosi etape - juvenaliniame - miško atsikūrimo ir bendrijų susidarymo ekologiniu
požiūriu rizikingiausiose - gamtinių ir antropogeninių trikdžių suardytose miško ekosistemose.
Mūsų vykdomais tyrimais yra siekiama išsiaiškinti, kaip klimato kaita ir kiti aplinkos
streso veiksniai veikia naujai besiformuojančias miško ekosistemas, jų produktyvumą ir biologinę įvairovę,
kokie yra miško ekosistemų raidos, produktyvumo, konkurencingumo ir biologinės įvairovės pokyčių dėsningumai kintant klimatui.
Tam tiriamas klimato kaitos ir kitų veiksnių – didėjančios temperatūros, drėgmės ir CO2 koncentracijos,
dažnėjančių sausrų, kaitros ir šalčio bangų, didėjančios pažemio ozono koncentracijos,
intensyvėjančios UV spinduliuotės – kompleksinį poveikį atsikuriančių miško medžių bendrijų
augimui, struktūrai, populiacijų ir individų būklei. Siekiama naujų
žinių apie glaudžiai sąveikaujančių skirtingų rūšių tarpusavio santykius, galimus
medžių konkurencingumo pokyčius ir to įtaką besiformuojančių miško bendrijų rūšinei sudėčiai
ir biologinei įvairovei. Svarbūs kompleksiniai medžių prieaugio, jų sezoninio vystymosi ir būklės bei tai lemiančių
fiziologinių procesų priklausomybės nuo klimato ir aplinkos pokyčių tyrimai.
Svarbu įvertinti ekosistemų gebėjimą adaptuotis prie kintančių klimato ir kitų aplinkos sąlygų.
Taip pat svarbu ištirti pagrindinių medžių rūšių genetinę įvairovę rizikingiausiose trikdžių paveiktose
buveinėse ir įvertinant galimą jos įtaka bendrijų ir populiacijų tvarumui
bei gebėjimui prisitaikyti (adaptyvumui). Taip pat svarbu atskleisti apleistose žemėse
savaime atželiančių miškų biologinės ir genetinės įvairovės formavimosi dėsningumus,
įvertinti šių naujų ekosistemų tvarumą bei gebėjimą prisitaikyti prie kintančių sąlygų.
Šie nacionalinės programos projekto tyrimai atliekami skirtingų mokslo šakų sandūroje:
miškotyros, augalų ekologijos, fiziologijos, genetikos ir fitopatologijos.
Tyrimai turėtų suteikti naujų žinių apie skirtingų bioekologinių savybių
medžių rūšių ir jų besiformuojančių bendrijų bei populiacijų eko-atsaką ir plastiškumą sąryšyje
su aplinkos sąlygų stresiškumu, augimo ritmu skirtingomis gamtinėmis ir fitotrone sukuriamomis
kompleksinėmis stresinėmis aplinkos sąlygomis, nustatant labiausiai lemiančias;
nustatyti miško bendrijų atsikūrimo ir formavimosi dinamiką klimato kaitos ir kitų stresorių
poveikyje ekologiškai rizikingiausiose trikdžių paveiktose miško ekosistemose.
Tyrimų rezultatų pagrindu būtų parengtos miško bendrijų vystymosi prognozės ir
rekomendacijos miško ekosistemų būklei stabilizuoti, joms atkurti, atsparumui stresams
didinti ir tvarumui užtikrinti. Tyrimų rezultatai bus pateikti ASU universitetinių studijų programoms
"Miško ekosistemos" ir "Miškininkystė"> (studijų dalykas 'Miško ekosistemų dinaminis
tvarumas") - jų dėstytojams ir klausytojams kaip metodinė ir vaizdinė problematikos medžiaga.
Projekto tikslas - ištirti septynių ūkiniu požiūriu svarbiausių miško medžių rūšių
ir jų populiacijų atsaką, plastiškumą ir konkurencijos pokyčius padidintos temperatūros, drėgmės
ir CO2 koncentracijos sąlygomis ir stresinių veiksnių - šalnų, karščio bangų, sausrų, padidinto
UV intensyvumo ir ozono koncentracijos kompleksiniame poveikyje miško bendrijų atsikūrimo ir
formavimosi stadijoje ir tuo pagrindu parengti rekomendacijas miško ekosistemų tvarumui užtikrinti.
Tyrimų uždaviniai:
1. Nustatyti skirtingų medžių rūšių - paprastojo ąžuolo, paprastojo uosio, paprastosios pušies,
paprastosios eglės, karpotojo beržo, juodalksnio ir drebulės ir jų populiacijų atsaką ir
plastiškumą pagal kompleksą rodiklių - augimo spartą, sezoninio vystymosi ritmą (fenologiją),
fiziologinius parametrus, sanitarinę būklę (ligų pažeidimus) ir kt. su klimato kaita susijusių
abiotinių veiksnių kompleksiniame poveikyje klimatinėse kamerose;
2. Ištirti atskirų medžių rūšių vidurūšinės ir tarprūšinės konkurencijos pokyčius ir to
pasekmes individualiam ir bendrijų vystymuisi, augimui ir rūšinei įvairovei su klimato kaita
susijusių abiotinių veiksnių kompleksiniame poveikyje, imituojant sudėtinių miško bendrijų
kūrimąsi klimatinėse kamerose;
3. Ištirti ir palyginti skirtingų medžių rūšių natūraliai besiformuojančių miško bendrijų
augimo, konkurencijos ir biologinės įvairovės specifiką gamtinių stresorių poveikyje ekologiškai
rizikingiausiose skirtingų trikdžių paveiktose gamtinėse buveinėse - vėjovartų, kirtaviečių ir
ligų suardytose miško bendrijose bei apleistuose žemės ūkio plotuose, gautus duomenis palyginant
su klimatinėse kamerose gautais rezultatais;
4. DNR mikrosatelitinių žymenų metodu ištirti atsikuriančių ir besiformuojančių miško bendrijų
karkasinių medžių rūšių genetinę įvairovę ekologiniu požiūriu rizikingiausiose trikdžių paveiktose
kertinėse gamtinėse buveinėse įvertinant jos pakankamumą tvarioms populiacijoms suformuot;
5. Nustatyti skirtingų dirbtinių ir gamtinių adaptacinių aplinkų selektyvumą, atskleidžiantį rūšių,
jų formuojamų bendrijų bei populiacijų skirtumus pagal atsaką, plastiškumą, konkurentingumą
ir genetinę įvairovę;
6. Parengti miškininkystės praktikos adaptavimo rekomendacijas miško ekosistemų tvarumui užtikrint;
I. Skirtingų medžių rūšių ir populiacijų ekoatsakas ir plastiškumas su klimato kaita susijusių
abiotinių veiksnių kompleksiniame poveikyje tiriami imituojant
miško medžių bendrijų formavimąsi dirbtinėmis aplinkos sąlygomis LAMMC Miškų instituto Fitotrono klimatinėse kamerose (1 pav.),
taikant 4 kompleksinio poveikio variantus: a) kontrolinis - lauko daigyno sąlygomis;
b) padidintos temperatūros (T), drėgmės (Dr) ir CO2 koncentracijos sąlygomis;
c) padidintos T, Dr ir CO2 sąlygomis ir stresinių veiksnių - šalnų, karščio bangų ir sausrų poveikyje;
d) padidintos T, Dr ir CO2 sąlygomis ir stresinių veiksnių - padidintų UV
spinduliuotės intensyvumo ir ozono koncentracijos poveikyje;
e) padidintos T, Dr ir CO2 sąlygomis ir stresinių veiksnių šalnų, karščio bangų,
sausrų, padidintų UV spinduliuotės intensyvumo ir ozono koncentracijos poveikyje.
Šiose klimatinėse kamerose konteineriuose auginami ir testuojami 2-4 m. amžiaus paprastojo ąžuolo (Ą), papr. uosio (U),
papr. pušies (P), papr. eglės (E), karpotojo beržo (B), juodalksnio (J) ir drebulės (D) sodinukai, atstovaujantys po tris
populiacijas iš skirtingų Lietuvos miško gamtinių regionų. Sudarytos 7 vienarūšės ir 3 mišrios dirbtinės juvenalinės medžių
bendrijos (po 500 vnt., iš viso 5000 medelių) (1 pav.).
Atsakas ir plastiškumas tiriamas pagal kompleksą rodiklių -
augimo spartą, sezoninio vystymosi ritmą (fenologiją), įvairius fiziologinius ir biocheminius parametrus, ligų pažeidimus
ir kt.
1 pav. Pagrindinių medžių rūšių ir jų populiacijų palikuonių bandymai LAMMC Miškų instituto Fitotrono klimatinėse kamerose.
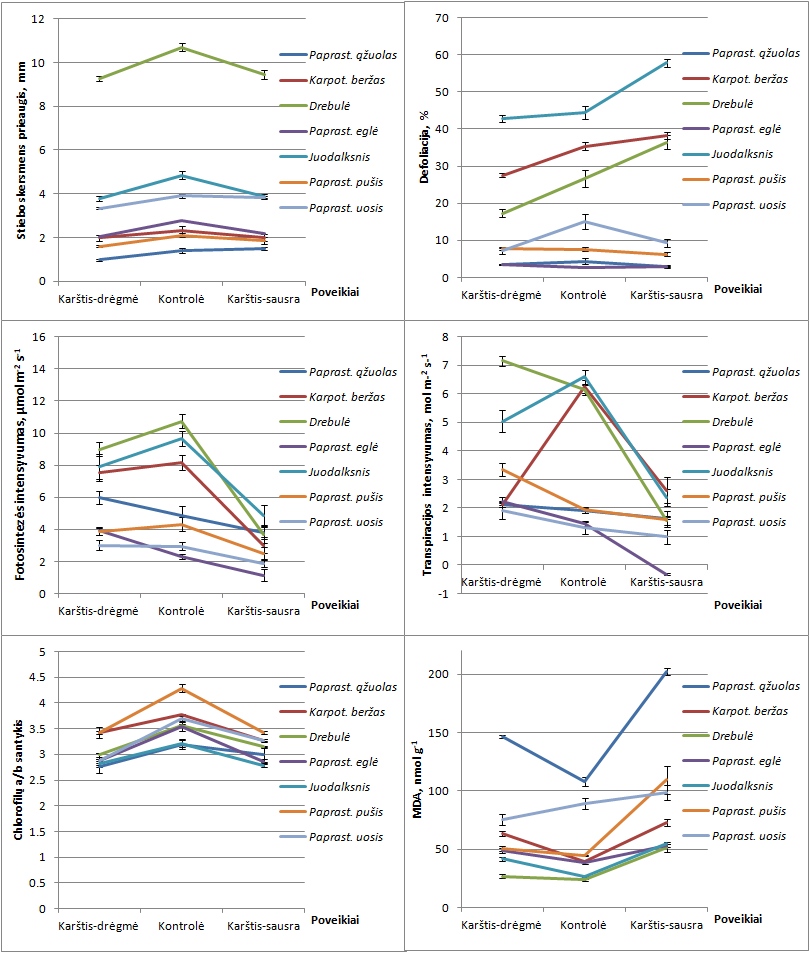
Nustatyta patikima poveikių karščiu-drėgme ir karščiu-sausra įtaka medžių augimui, fiziologiniams ir
biocheminiams rodikliams – aukščio ir skersmens prieaugiui, aukščio ir skersmens santykiui, defoliacijai,
išlikimui, žiotelių pralaidumui, vidiniam vandens naudojimo efektyvumui (WUE), chlorofilo a ir b kiekiui
bei chlorofilo a/b santykiui, karotenoidų, MDA ir H2O2 kiekiui lapuose ir spygliuose.
Tiek karštis-drėgmė, tiek karštis-sausra labiau neigiamai atsiliepė medžių skersmeniui nei aukščiui,
tad pakito ir aukščio ir skersmens santykis.
Defoliacija reikšmingai padidėjo karščio-sausros sąlygomis,
ypač pionierinių lapuočių medžių rūšių – drebulės, juodalksnio ir karpotojo beržo
(atitinkamai iki 36,4±1,8, 57,8±1,1 ir 38,5±0.9%),
o sumažėjo karščio-drėgmės sąlygomis. Chlorofilo a ir b bei karotenoidų kiekis
padidėjo karščio-drėgmės poveikyje visoms rūšims (išskyrus eglės).
Labiau stresinėmis sąlygomis – karščio-sausros poveikyje
– daugelyje tirtų medžių rūšių chlorofilo a ir b bei karotenoidų kiekis reikšmingai sumažėjo.
Chlorofilo a/b santykis sumažėjo tiek karščio-sausros, tiek ir karščio-drėgmės poveikyje.
Taigi, stresinės sąlygos turėjo neigiamos įtakos pigmentų gamybai ir fotosintezei, tačiau atskirų medžių
rūšių reakcija buvo labai skirtinga. MDA koncentracija, indikuojanti augalų oksidacinį stresą,
daugeliui tirtų medžių rūšių padidėjo dėl abiejų stresinių poveikių. H2O2 koncentracija sumažėjo dėl
karščio-drėgmės poveikio, o dėl karščio-sausros poveikio ji kito priklausomai nuo medžių rūšies.
Dendrometrinių, fiziologinių ir biocheminių rodiklių pokyčiai indikuoja nemažą medžių fenotipinį
plastiškumą, tačiau išlieka neaišku, ar jis yra adaptacinio pobūdžio, ar tik atspindi medžių fiziologinės būklės pablogėjimą.
Nustatyta reikšminga rūšių ir jų sąveikos su poveikiu įtaka visiems tirtiems požymiams rodanti,
jog tirtos rūšys pasižymi specifiniu biocheminiu, fiziologiniu, morfologiniu ir prieaugio atsaku į
kompleksinį stresorių poveikį – kiekviena jų pasižymėjo savito pobūdžio ir
stiprumo ekoatsaku (2 pav.).
2 pav. Pagrindinių medžių rūšių atsakas į stresines sąlygas pagal skirtingus požymius.
Rūšių atsakas labiausiai skirtingas buvo pagal chlorofilo a ir b kiekio, transpiracijos, MDA
ir H2O2 koncentracijų pokyčius po skirtingų poveikių. Po karščio-sausros poveikio daugelio medžių
rūšių aukščio ir skersmens prieaugiai lyginant su kontrole sumažėjo, o paprastojo ąžuolo ir
juodalksnio aukščio prieaugiai po karščio-drėgmės ir karščio-sausros net padidėjo (2 pav.),
kas gali padidinti šių rūšių konkurencingumą šiltėjant klimatui
Drebulė ir karpotasis beržas dėl karščio-sausros nukentėjo labiausiai – drebulės fotosintezė
sumažėjo 66 %, beržo – 63 %, juodalksnio – 50 %, dėl ko labiausiai sumažėjo jų aukščio prieaugis.
Kaip indikuoja karščio-sausros poveikyje padidėjęs H2O2 juodalksnio ir uosio lapuose ir pušies bei eglės spygliuose,
šios rūšys yra jautriausios sausros stresui. Tos medžių rūšys, kurios pasižymėjo didžiausia transpiracija
įprastomis sąlygomis – juodalksnis, beržas ir drebulė, pasižymėjo savita reakcija į sausrą –
jos karščio-sausros sąlygomis ypač stipriai sumažino transpiraciją bei numetė daug lapų.
Kartu labai sumažėjo fotosintezės intensyvumas. Po karščio-drėgmės poveikio transpiracijos sumažėjimas užfiksuotas
tik juodalksniui ir beržui, o kitoms medžių rūšims transpiracija padidėjo. Skirtumai tarp rūšių pagal WUE
buvo nedideli įprastomis sąlygomis, tačiau karščio-sausros poveikyje lyginant tiek su įprastomis sąlygomis,
tiek ir su karščio-drėgmės sąlygomis paprastosios pušies, paprastosios eglės ir karpotojo beržo WUE sumažėjo,
o drebulės, juodalksnio ir paprastojo ąžuolo – padidėjo.
Nustatytas reikšmingas populiacinis efektas daugeliui požymių, išskyrus transpiracijos intensyvumui,
žiotelių pralaidumui ir WUE, kurių populiacinis efektas buvo mažiau reikšmingas arba nereikšmingas.
Pastariesiems ir daugumai kitų požymių (išskyrus išsilaikymą ir medžių būklę) labai reikšmingas buvo populiacijų
ir bandymų sąveikos efektas (Gp x Eb), rodantis, kad populiacijos pasižymi specifiniu atsaku į tirtų stresinių sąlygų poveikį.
Vertinant kiekvienos populiacijos plastiškumą ir ekovalentingumą pagal aukščio prieaugį buvo nustatyta,
kad tik 4 iš tirtos 21 populiacijos labiau keitė rangus po skirtingų bandymų ir esminiai prisidėjo prie GxE
sąveikos su 11,1-21,4% ekovalentingumu.
Transpiracijos, žiotelių pralaidumo ir fotosintezės intensyvumo tarppopuliacinė variacija labai skyrėsi
skirtingomis bandymų sąlygomis ir kontrolėje: didžiausias tarppopuliacinės variacijos koeficientas po
karščio-sausros bandymo nustatyta pušiai, eglei, juodalksniui ir drebulei, uosiui - po karščio-drėgmės bandymo,
o ąžuolo - kontrolinėmis sąlygomis. Ypač didelė WUE tarppopuliacinė variacija daugeliui medžių rūšių
(išskyrus uosio) nustatyta labiausiai stresiniame karščio-sausros bandyme. Labiausiai išreikšta Gp x Eb
sąveika nustatyta chlorofilo a ir b koncentracijoms bei jų santykiui a/b, karotenoidų, MDA ir H2O2 koncentracijoms.
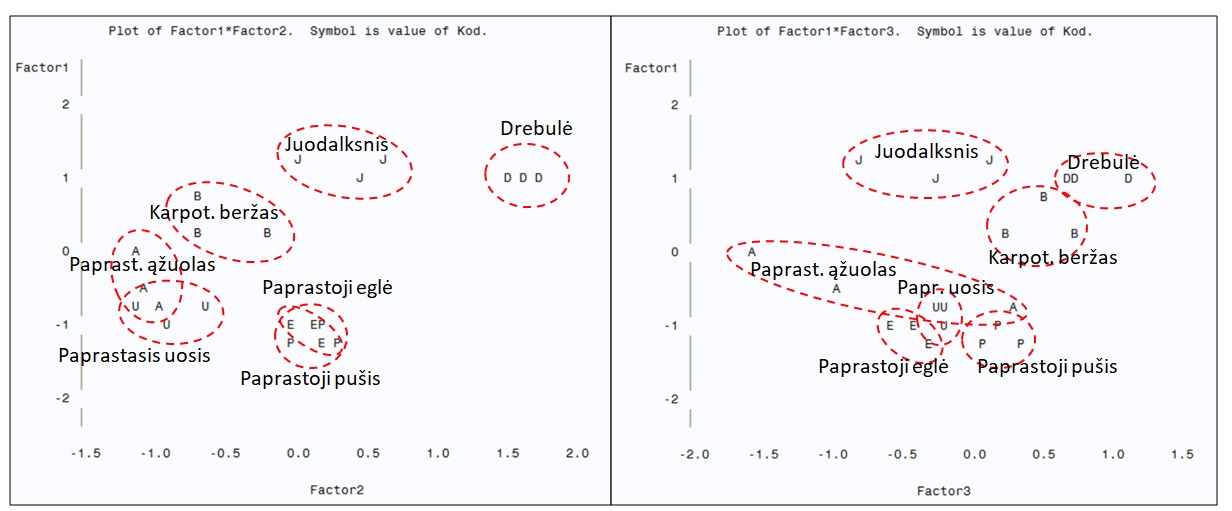
Visų tirtų populiacijų visų požymių daugiamatė faktorinė analizė atskleidė tris pagrindinius faktorius.
Pirmasis faktorius labiausiai koreliavo su karotenoidų, chlorofilo a ir b koncentracijomis, jų santykiu
a/b, H/D santykiu, defoliacija, transpiracija ir fotosinteze, antrasis faktorius teigiamai koreliavo su
aukščio ir skersmens prieaugiais, neigiamai – su MDA ir H2O2 koncentracijomis, o trečiasis – su chlorofilo
a/b santykiu, fotosintezės intensyvumu ir H2O2 koncentracija. Pagal daugiamatę analizę labiausiai tarpusavyje
ir nuo kitų rūšių išsiskyrė drebulės, juodalksnio ir beržo populiacijos, kas rodo jų ekologinės elgsenos
išskirtinumus (3 pav.). Pušies ir eglės populiacijos susigrupavo arti vienos kitų pagal pirmąjį faktorių.
Ąžuolo ir uosio populiacijos susigrupavo pagal pirmąjį ir antrąjį faktorius, kas rodo jų panašią ekologinę elgseną.
3 pav. Pagrindinių medžių rūšių ir populiacijų išsidėstymas pagrindinių komponenčių erdvėje
pagal komplekso požymių išreikštumą skirtingomis bandymų sąlygomis.
II. Medžių vidurūšinės ir tarprūšinės konkurencijos pokyčių ir jų pasekmių individualiam ir bendrijų
vystymuisi, augimui ir rūšinei įvairovei tyrimai vsu klimato kaita susijusių abiotinių veiksnių
kompleksinio poveikio sąlygomis (4 kompleksai) Fitotrono klimatinėse kamerose (4 Pav.),
Imituojamas sudėtinių miško bendrijų kūrimąsis, tam sudarant dirbtines tiriamų medžių bendrijas - skirtingus
mišinius, charakteringus trims miško augaviečių tipams:
a) Nb - P+B+E, b) Lc - E+B+Ą, c) Lf - U+J+D.
Matuojami augimo, sezoninio vystymosi ritmo, fiziologiniai, biocheminiai, sanitarinė būklės, biomasės pasiskirstymo
ir kt. rodikliai ir tiriama, kaip jie keičiasi priklausomai nuo mišinio rūšinės sudėties, šalia augančių medžių rūšių
ir skirtingų aplinkos sąlygų skirtingose kamerose. Šiuo metu tyrimų duomenys analizuojami ir rengiamas mokslinis straipsnis į tarptautinį mokslo žurnalą.
4 pav. Pagrindinių medžių rūšių mišinių bandymai karščio-drėgmės sąlygomis.
5 pav. Pagrindinių medžių rūšių mišinių bandymai karščio-sausros sąlygomis.
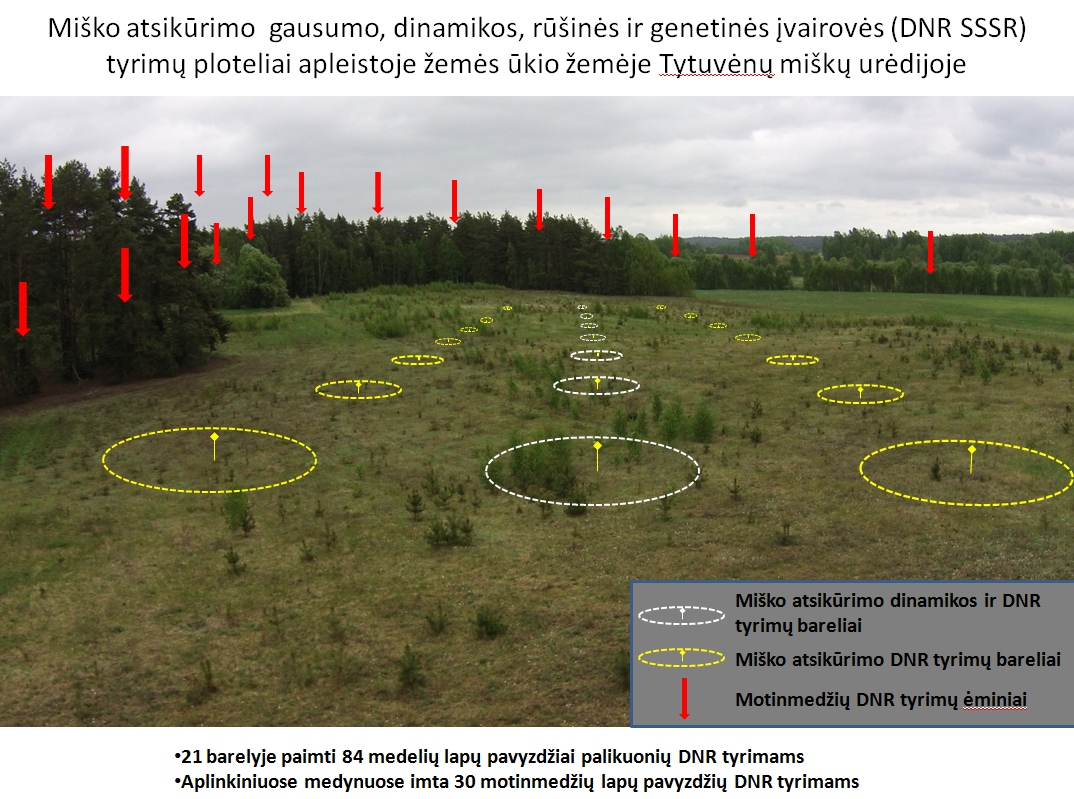
III. Natūraliai atsikuriančių medžių bendrijų atsikūrimo tyrimai atliekami
Ą, U, P, E, B, J savaiminiuose 3-7 metų amžiaus žėliniuose skirtingų
trikdžių paveiktose rizikingiausiose miško gamtinėse buveinėse:
a) vėjo sudarkytose (E, P), b) ligų ir kenkėjų pažeistose (U, E), c) plynose kirtavietėse (E, P, J),
ir d) dirvonuojančiuose žemės ūkio plotuose (P, B). 2015 metais
Dubravos, Tytuvėnų, Anykščių, Kėdainių, Prienų urėdijų miškuose kiekvienoje tipinėje buveinėje išskirta
po 2-3 tyrimo plotus po 6-8 barelius - uždėta 16 serijų ilgalaikių barelių (iš viso 112 barelių),
kuriuose kasmet vegetacijos zezono gale registruojami ir matuojami visi medžių sėjinukai, žolinė
danga, trakas, dirvožemio charakteristikos, aplinkiniai motinmedžiai ir kt. (6 pav.)
6 pav. Atsikūrimo tyrimų bareliai pušimi ir beržu atsikuriančiame apleistame žemės ūkio
naudmenų plote Tytuvėnų miškų urėdijoje
Tyrimai parodė, jog rizikingiausiose gamtinių trikdžių – vėjovartų ir ligų bei kirtimų ir žemės ūkio
veiklos sutrikdytose miško ekosistemose, miško atsikūrimas vyksta nepriklausomai nuo trikdžio tipo.
Didelė atsikūrimo kiekybinių ir kokybinių rodiklių variacija rodo, kad kuriasi platus spektras naujų
bendrijų, kurios dažniausiai skiriasi nuo anksčiau buvusių. Rūšių sukcesija pasireiškia jau
ankstyvoje bendrijų formavimosi stadijoje. Atsikuriančių bendrijų rūšinė sudėtis priklauso nuo buvusių
prieš trikdžius, tačiau sutrikdytoms ekosistemoms būdingos pionierinės rūšys – beržai, pušys,
juodalksniai ir kt. pradeda dominuoti.
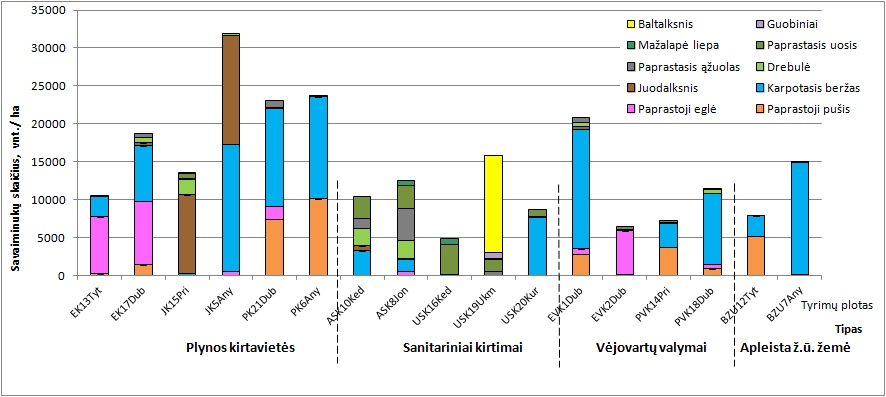
Intensyviausias miško atsikūrimas vyksta tose ekosistemose, kurios paveiktos tiesioginiu žmogaus
poveikiu – plynose kirtavietėse, o mažiau intensyvus, ypač tikslinėmis medžių rūšimis – vėjovartų
ir sanitarinių kirtimų plotuose (7 pav.). Nors vyrauja ne tikslinių konkrečiai augavietei medžių rūšių
savaiminukai, tačiau tikslinių rūšių savaiminukų kiekis yra pakankamas produktyvioms miško
ekosistemoms atsikurti ar naujoms susidaryti. Tikslinių rūšių atsikūrimo gausumas palankus
biologinei įvairovei ir rūšies išlikimui, tačiau siekiant suformuoti produktyvius miškus, reikalinga
miškininkystės priemonėmis kontroliuoti konkuruojančių pionierinių medžių ir žolinės augalijos
rūšių gausumą.
7 pav. Skirtingų medžių rūšių savaiminukų kiekiai skirtingose sutrikdytose miško ekosistemose.
Daugiamatė regresinė analizė parodė, jog skirtingų medžių rūšių atsikūrimo gausą lemia skirtingas kompleksas
6-8 vietinės aplinkos biotinių ir abiotinių veiksnių, dažniausiai turinčių neigiamą poveikį
atsikūrimui: atstumas iki motinmedžio, didelis buvusio medyno skalsumas, didelis padengimas
negyva mediena, lajomis, traku, atskiromis trako ir žolių rūšimis ir kt.
Nustatyta didelė atsikūrimo gausumo, jo erdvinio tolygumo, augimo diferenciacijos, rūšių įvairovės
ir susimaišymo variacija atsikuriančiuose plotuose (8 pav.), kurią sąlygoja stipri dėmių struktūros
edafinių sąlygų bei žolinės dangos variacija, intensyvėjanti dėl klimato kaitos dažnėjančių
temperatūrinių ir drėgmės ekstremumų, būdingų trikdžių paveiktuose atviruose plotuose.
Biologinės įvairovės indeksų analizė atskleidė, jog labiausiai sutrikdytose ekosistemose
dominuoja tik viena ar dvi medžių rūšys, kurios formuoja naujojo medyno pagrindą, tačiau
plačialapių rūšių medynuose, kertamuose sanitariniais kirtimais, randama didesnė medžių
rūšių gausa be atskirų rūšių dominavimo.
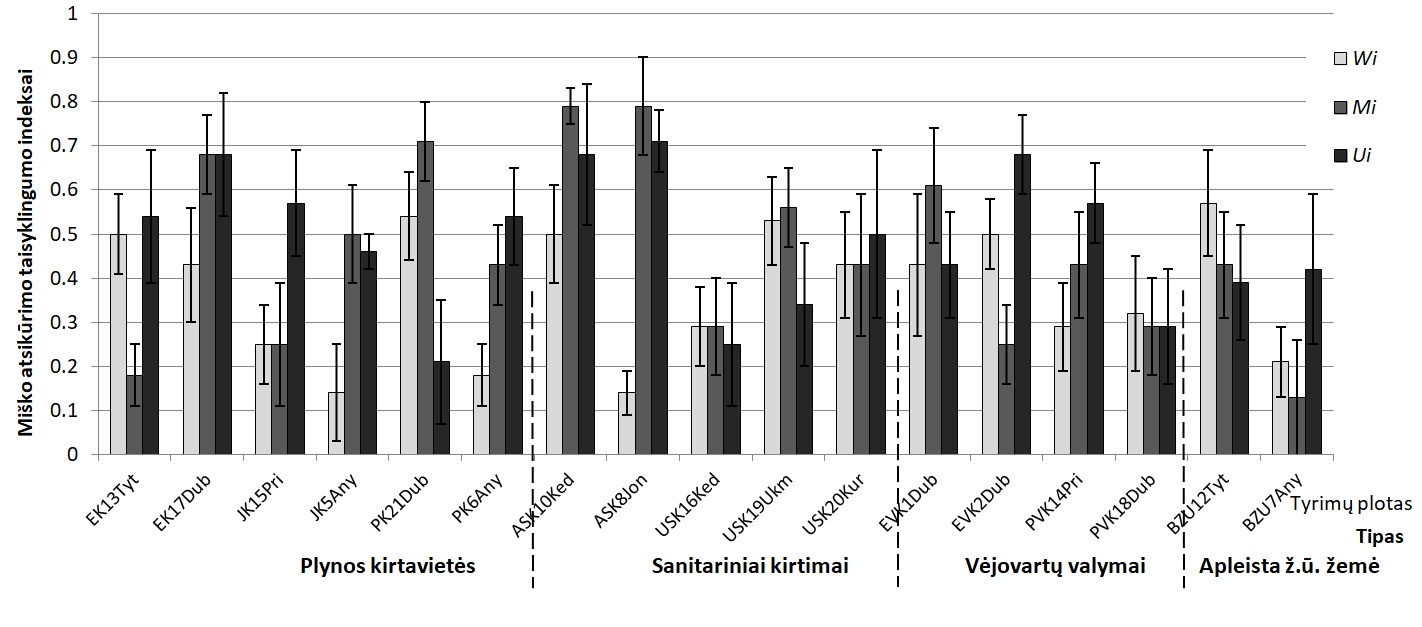
8 pav. Atsikūrimo tolygumo indeksai skirtingose sutrikdytose miško ekosistemose. Wi - medelių
išsidėstymo tolygumo indeksas, Mi - rūšių susimaišymo indeksas, Ui - dominavimo indeksas .
IV. Atsikuriančių miško bendrijų karkasinių medžių rūšių genetinė įvairovė tiriama
ekologiniu požiūriu rizikingiausiose trikdžių paveiktose ktipinėse buveinėse.
Kiekviename iš 16 tyrimų plotų imami savaiminukų lapų ar spyglių
pavyzdžai 7-se tyrimo bareliuose bei papildomuose 14-je barelių (84 iš viso pavyzdžiai plote) (2 Pav.). Be to,
palyginimui su ankstesnės generacijos genetine įvairove, imami pavyzdžiai nuo aplinkiniuose
besiribojančiuose sklypuose augančių galimų motinmedžių (20-30 medžių).
Savaime gausiai atsikuriančios eglės plynose kirtavietėse, o pušys ir beržai - apleistuose žemės
ūkio plotuose nustatyta didele genetinė įvairovė, kuri netgi viršija aplinkinių motininių medynų
įvairovę, tad šių miško ekosistemų atsikūrimo genetiniam nepilnavertiškumui pavojaus nėra.
Eglės ir beržo palikuonių ir gretimų motininių medynų individų persidengiantis išsidėstymas SSR
žymenų Principinių komponenčių erdvėje (9 pav.), maži Nei genetiniai atstumai ir maži genetinės variacijos
tarp medynų ir žėlinių skirtumai rodo, kad plotuose, kuriuose vyksta gausus atsiželdymas, genetinė
įvairovė lyginant su kaimyniniais medynais nesumažėja, palikuonys yra kilę iš šių medynų, tačiau
pušys, atsikuriančios apleistuose žemės ūkio plotuose, yra genetiškai labiau skirtingos nuo
kaimyninių motininių medynų dėl tolimesnės genų pernašos.
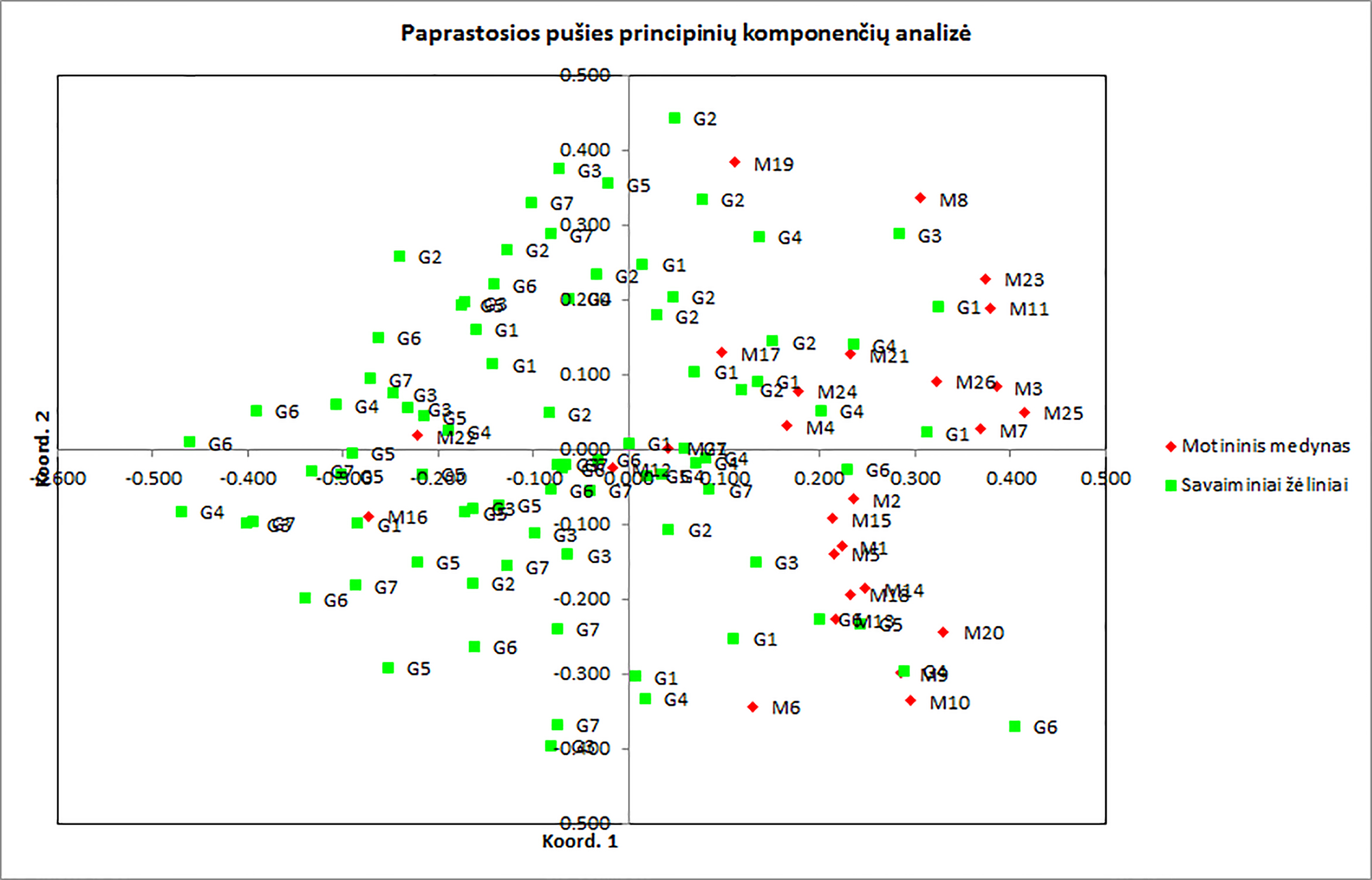
9 pav. Paprastosios pušies individų iš motininio medyno (M) ir iš savaiminių žėlinių (G) trikdžių paveiktuose
plotuose DNR SSR molekulinių žymenų PCA analizės rezultatai. Pirmosios dvi komponentės paaiškina 44,61% variacijos
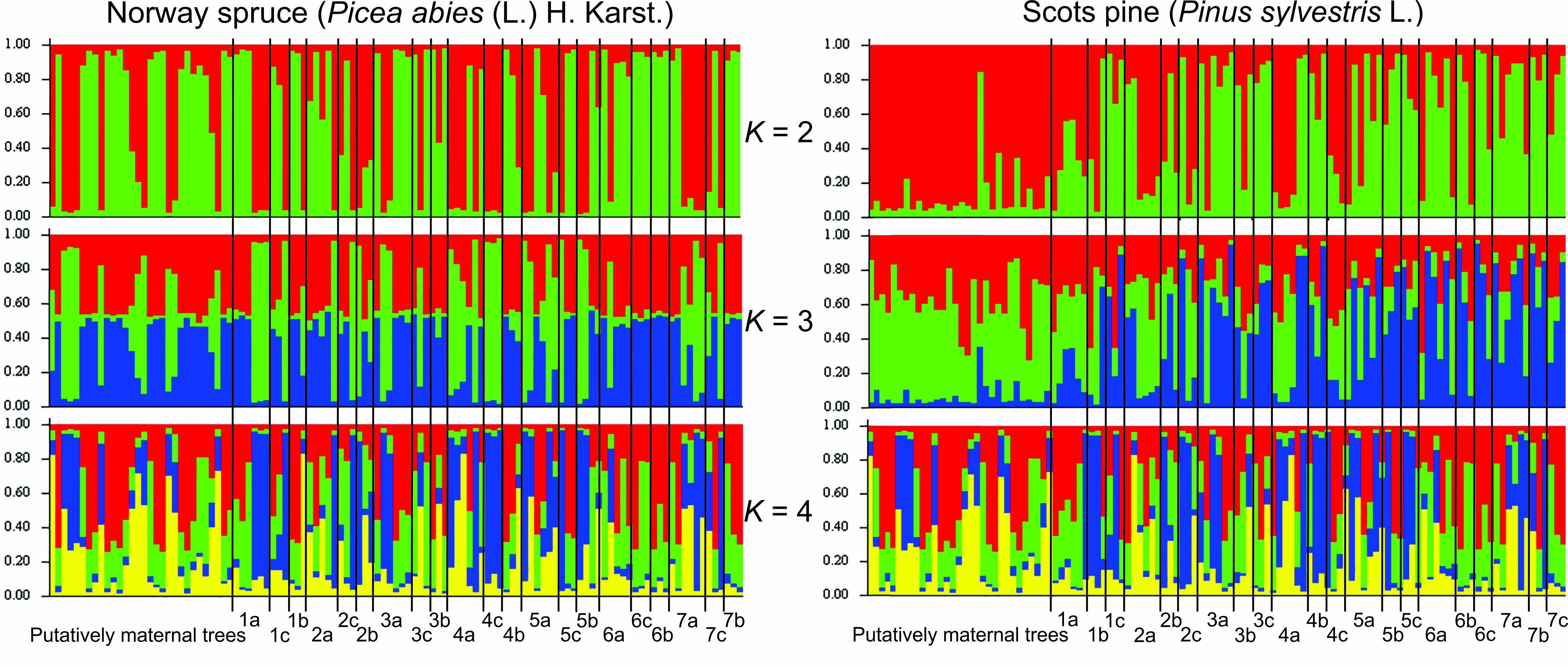
Pušies ir eglės atsikūrimo genetinę struktūrą patikimai atsispindi trys genetiniai klasteriai (10 pav.),
o beržo – keturi. Tačiau pušies motinmedžių struktūra paprastesnė - nustatyti tik du genetiniai
klasteriai, kas rodo, jog dalis palikuonių yra kilę iš genetiškai skirtingesnių medžių, kurie randasi
tolimesniuose medynuose.
10 pav. Paprastosios eglės ir pušies medynų (motinmedžiai) ir žėlinių (grupės G1-G7)
DNR SSR žymenų klasterinė genetinė struktūra
Erdvinės genetinės struktūros analizė nei vienai iš trijų tirtų rūšių neatskleidė ryškesnės
erdvinės struktūros, kas yra palanku atsitiktiniam kryžminimuisi (11 pav.). Artimi savaiminukai grupėse
nesudaro atskirų genetiškai giminingų grupių ir tik atskirais atvejais grupėse 2-3 kaimyniniai
medžiai yra giminingi.
11 pav. Paprastosios pušies savaiminukų, priskiriamų skirtingiems genetiniams klasteriams
(pažymėta baltai, pilkai ir juodai), pasiskirstymas
saviminiuose žėliniuose apleistose žemės ūkio žemės plotuose.
Išsamūs tyrimų rezultatai pateikti straipsniuose tarptautiniuose mokslo žurnaluose:
Pliūra A., Jankauskienė J., Lygis V., Suchockas V., Bajerkevičienė G., Verbylaitė R.
Response of juvenile progeny of different populations of seven forest tree species to simulated
climate change-related stressors, heat and drought. iForest - Biosciences and Forestry (IF=1,62)
Verbylaitė R., Pliūra A., Lygis V., Jankauskienė J., Labokas J. Genetic diversity and its
spatial distribution in self-regenerating Norway spruce and Scots pine stands. Forests (IF=1,9)
Suchockas V., Pliūra A., Labokas J., Lygis V., Dobrowolska D., Jankauskienė J., Verbylaitė R.
Evaluation of early stage regeneration of forest communities following natural and human-caused
disturbances. Forestry: An International Journal of Forest Research (IF=2,2)
Tyrimų rezultatų dalis pristatyta tarptautinėje IUFRO konferencijoje Tokijuje, Japonija, 2017 spalio 22-26 d.:
Padarytas pranešimas Pliūra A.*, Suchockas V., Jankauskienė J., Lygis V., Verbylaitė R., Labokas J.
"Response of seven forest tree species to simulated climate change stressors, heat and drought".
Santrauka išspausdinta konferencijos leidiniyje Proc. IUFRO conference Forest ecosystems under air
pollution and climate change, 22-26 October,2017, Tokyo University of Agriculture and Technology,
Tokyo, Japan, p. 40.
LAMMC, LAMMC, LAMMC, LAMMC, LAMMC,
LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC
LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC, LAMMC




Lietuvos miško genetinių išteklių svetainė ![]()
![]() Atgal
Atgal